Toda
a água que passa pela planta provém do solo. A demanda
evaporativa da atmosfera provoca uma perda da água pelo
vegetal ocasionando uma necessidade contínua de reposição
da água perdida que se estende desde as folhas até
as raízes passando pelo xilema (Figura 8).
Nos vasos xilemáticos ocorre a formação de
uma coluna capilar de água sob tensão que se estende
até as folhas. A coluna de água somente se mantém
intacta, mesmo sob elevadas tensões, graças à
coesão entre as moléculas de água. Essa tensão
xilemática se transfere até as raízes, provocando
um gradiente de potencial hídrico entre raiz e solo, favorecendo
o fluxo em massa ou osmose da água, do solo para o interior
das raízes.
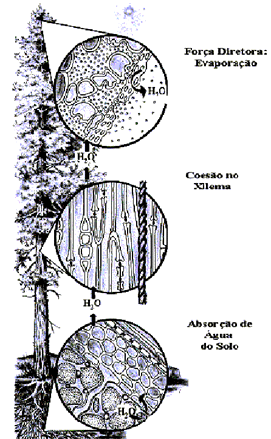
Figura 8.
Resumo do fluxo em massa de água no sistema solo-planta-atmosfera.
São geradas tensões foliares resultantes da transpiração,
devido ao gradiente de pressão de vapor entre a folha
e a atmosfera. Assim, uma coluna capilar de água sob
tensão se movimenta para as folhas. A coluna de água
é mantida intacta, mesmo sob elevadas tensões,
devido à coesão entre as moléculas de água.
Essa tensão xilemática se transfere até
as raízes, provocando um gradiente de potencial hídrico
entre raiz e solo, favorecendo o fluxo em massa ou osmose da
água, do solo para o interior das raízes.
Nas vizinhanças
das raízes o 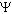 ws,
torna-se mais negativo (ponto A) sendo criado um gradiente de
potencial hídrico (
ws,
torna-se mais negativo (ponto A) sendo criado um gradiente de
potencial hídrico (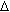 w)
em relação a um ponto B. Assim a água caminha
de A em direção a B percorrendo uma distancia
X (Figura 9).A
quantificação desse movimento, como citado anteriormente,
se faz utilizando a Equação de Darcy (Equação
7) na qual, w
representa a diferença de potencial hídrico entre
os pontos considerados, separados por uma distancia X (m) e
L, é a condutividade hidráulica do solo (m2 s-1
Pa-1).
w)
em relação a um ponto B. Assim a água caminha
de A em direção a B percorrendo uma distancia
X (Figura 9).A
quantificação desse movimento, como citado anteriormente,
se faz utilizando a Equação de Darcy (Equação
7) na qual, w
representa a diferença de potencial hídrico entre
os pontos considerados, separados por uma distancia X (m) e
L, é a condutividade hidráulica do solo (m2 s-1
Pa-1).
Jv = L (w
/x) Equação 7
Onde Jv,
representa o fluxo de água que é a quantidade
de água q (m3) que atravessa a unidade de área
transversal A (m2) na unidade de tempo t (s), ver Equação
7.1:
Jv = q / (A/t)
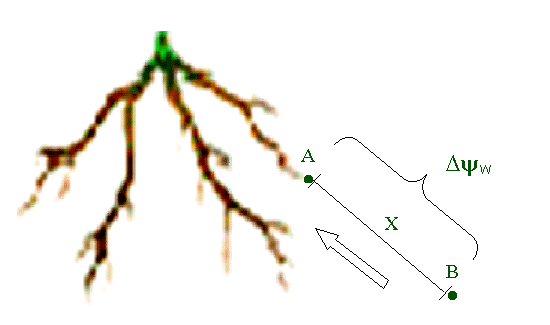
Figura 9.
Modelo esquemático demonstrando o movimento da água
no solo em direção a raiz. A força motriz
para o movimento é o gradiente de potencial hídrico
(w)
entre os dois pontos considerados (pontos A e B) que estão
separados por uma distância X. O potencial hídrico
em A é menor do que em B, supondo-se que a raiz esteja
absorvendo água. Assim, o fluxo de água ocorre
de B para A (seta).
Água
entra na planta pela raiz, cujo xilema tem continuidade com
o do caule. As células da endoderme, externamente ao
córtex, formam uma barreira impermeável ao fluxo
radial e transversal de água, devido a suberificação
das paredes celulares conhecida como estria de Caspari. As paredes
celulares paralelas à superfície da raiz geralmente
não são impregnadas com suberina. Portanto, não
oferecendo resistência ao fluxo de água. O movimento
da água pode ocorrer através de duas rotas, a
saber: a rota simplástica, ou seja, através dos
protoplasmas das células unidos pelo plasmodesmata ou
pela rota apoplástica movendo-se através das paredes
celulares e espaços intercelulares (Figura 10).
A penetração da água e solutos tende a
ocorrer mais rapidamente através das regiões que
oferecem menor resistência à sua movimentação.
Essas regiões variam de acordo com a espécie,
idade e velocidade de desenvolvimento da raiz.
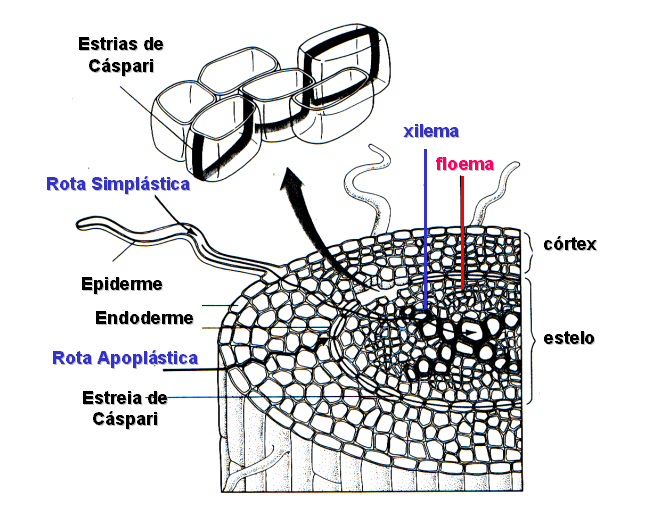
Figura 10. Modelo esquemático demonstrando o movimento
da água, após sua entrada na raiz, em direção
ao xilema. Note que o movimento da água pode ocorrer
através de duas rotas (simplásticas e apoplásticas)
até as células da endoderme que formam uma barreira
impermeável ao fluxo radial, transversal de água,
devido a suberificação das paredes celulares conhecida
como estrias de Caspari. Neste instante, a água atravessa
as membranas plasmáticas destas células e, conseqüentemente,
através da rota simplástica, alcança o
xilema, localizado no estelo.
A absorção
de água através da região meristemática
é bastante limitada, devido principalmente à grande
resistência oferecida pelo protoplasma denso e a falta
de elementos de condução nesta região.
Investigações anatômicas e fisiológicas
das regiões de absorção da raiz tem demonstrado
que a máxima absorção de água ocorre
na região de diferenciação celular onde
o xilema está bem diferenciado e na qual a suberização
e lignificação ainda não reduziram a permeabilidade
das paredes celulares.
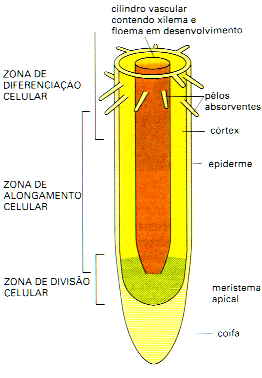
Figura 11.
Diagrama da extremidade de uma raiz mostrando quatro zonas ou
regiões distintas: coifa, região meristemática,
região de alongamento e de diferenciação
celular. Destas regiões, a máxima absorção
de água ocorre na região de diferenciação
celular devido tanto a maior diferenciação do
xilema quanto a incipiente suberização e lignificação
das paredes celulares
6.1 Forças
motrizes para o movimento de água
Para que ocorra a absorção de água, é
necessário o desenvolvimento de um gradiente de potencial
hídrico favorável no sistema solo-planta. Isto
se verifica quando há um aumento de potencial hídrico
na solução ao redor das raízes e/ou um
decréscimo do potencial hídrico nos vasos xilemáticos.
A velocidade na qual a absorção de água
se processa, depende da magnitude do gradiente do potencial
hídrico e das resistências oferecidas ao fluxo
de água no referido sistema.
A resistência ao movimento de água no solo depende
principalmente do conteúdo de água presente, enquanto
nas raízes resulta essencialmente, do grau de diferenciação
dos tecidos de condução, de suberização
e das condições físicas do protoplasma.
Outras resistências ao movimento da água também
podem ser citadas, tais como a resistência do xilema,
estomática, cuticular e da camada de ar limítrofe
(Figura 8).
O gradiente de potencial hídrico favorável ao
movimento da água para a atmosfera passando pela planta
pode resultar da ocorrência da transpiração
ou do acúmulo de íons no interior dos vasos do
xilema da raiz (pressão de raiz).
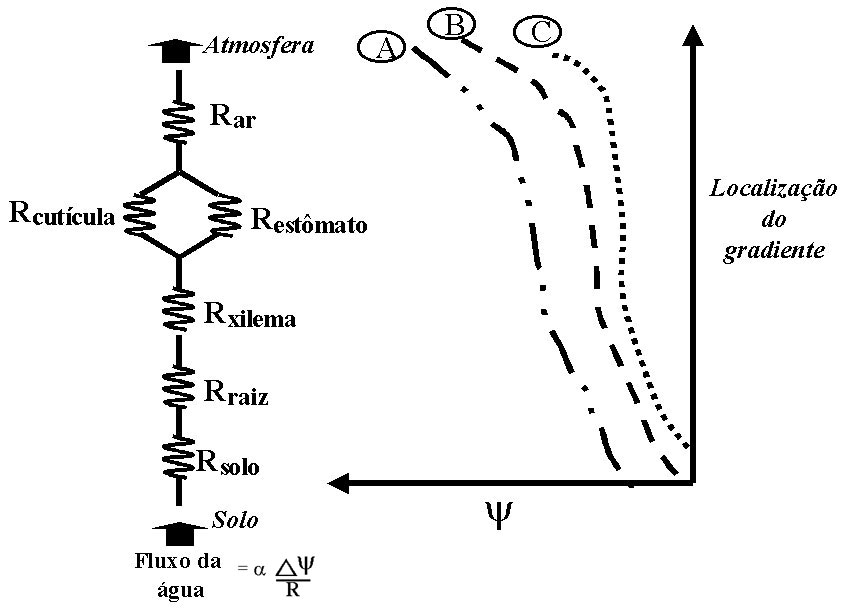
Figura 12. Potenciais hídricos e resistências ao
fluxo de água no sistema solo-planta-atmosfera, sob diferentes
condições: (A) dia normal; (B) durante a noite,
e (C) com solo seco.
6.1.1
Pressão de raiz
Embora a pressão de raiz não seja um fator, muito
importante para a manutenção do status hídrico
do vegetal, em determinadas condições como as
de alto potencial hídrico do solo e sob condições
de baixa transpiração, as raízes de muitas
plantas se comportam como osmômetros (ver Figura
4) desenvolvendo uma pressão positiva, chamada
pressão de raiz. A pressão de raiz é um
processo puramente osmótico e depende do acúmulo
de solutos no xilema, reduzindo seu potencial hídrico
para valores inferiores aos do substrato (solo). Portanto, a
água se movimenta desde o solo e penetra na raiz em função
de um gradiente de potencial hídrico crescente em virtude
principalmente do componente osmótico.
Porém, no simplasma e nos vasos xilemáticos da
raiz existiria um aumento crescente de íons o que dificultaria
o transporte lateral dos íons para dentro do simplasma
e, finalmente, para o interior dos vasos do xilema da raiz.
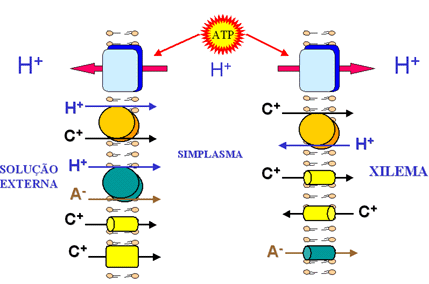
Figura
13. Modelo esquemático mostrando como os íons
são transportados contra seu gradiente de concentração
da solução externa para o xilema. As formas geométricas
representam os canais iônicos ou proteínas transportadoras
de cátions (C+) e ânions (A-) que atravessam as
membranas representadas pelas duas "redes de balões".
Observe que os quadrados superiores próximos ao ATP (adenosina
trifosfato) representam as proteínas ATPases que utilizam
a energia metabólica, na forma de ATP, para bombear prótons
(H+) que serão co-transportados ou antitransportados
com cátions ou ânions.
Como sempre
a concentração de íons no xilema é
superior a da solução externa, fica óbvio
a necessidade de um mecanismo de transporte ativo de íons,
mostrado na Figura 13. Por conseguinte, todo o transporte dependeria
da participação da energia metabólica,
na forma de ATP, e seria unidirecional visto que as membranas
são impermeáveis aos íons.
O acúmulo de íons no xilema induziria uma redução
do potencial osmótico, e por extensão, do potencial
hídrico, estabelecendo um gradiente favorável
ao movimento da água do solo para o interior do xilema.
O cilindro central ou estelo (ver Figura 10) passaria a funcionar
como um osmômetro perfeito, com as membranas que delimitam
o simplasto retendo seletivamente os íons ao mesmo tempo
as estrias de Caspari impediriam o retorno de sais pelo apoplasto.
Além da existência de proteínas canais e
transportadoras de íons e metabólitos, proteínas
denominadas aquaporinas seriam responsáveis pelo aumento
da permeabilidade à água nas raízes das
plantas pela formação de canais específicos
para a água por excluírem íons e metabólitos.
A entrada da água seguindo um gradiente favorável
causaria o desenvolvimento de uma pressão hidrostática
positiva e a ascensão da solução na forma
de um fluxo em massa denominada de pressão de raiz, resultando
na perda de água na forma líquida pela cutícula
foliar (gutação).
6.1.2
Transpiração
Outra força que define a ascensão da água
é a transpiração que depende do aparecimento
de um gradiente de potencial hídrico (?yw) favorável
más ocasionado pelo desenvolvimento de uma pressão
negativa ou tensão no interior dos vasos do xilema.
A força motriz adequada para gerar a tensão nos
vasos do xilema é a evaporação da água
das plantas (transpiração). Consideremos uma situação
na qual a planta se encontra com seus tecidos hidratados e há
disponibilidade de água no solo. Estando os estômatos
abertos e havendo uma demanda evaporativa, se verifica o processo
de transpiração.
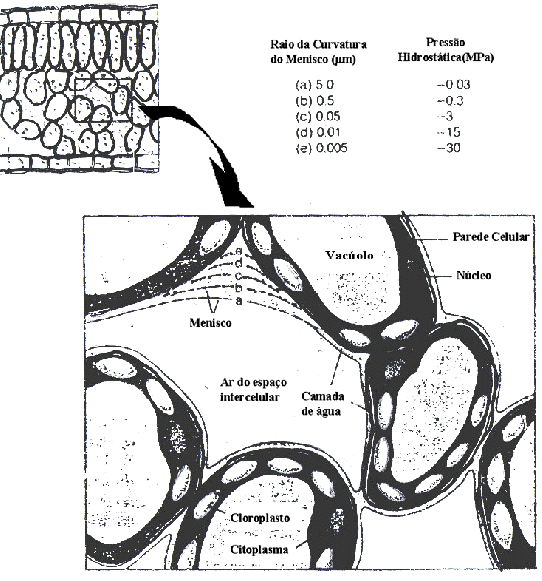
Figura
14. Esquema mostrando a origem das tensões (ou seja,
pressão negativa) na água presente na parede celular
de uma célula vegetal. Com a evaporação
da água que forme uma película sobre a parede,
meniscos são formados nos interstícios da parede,
os quais estão sob tensão. Quando o raio de curvatura
diminui, a pressão diminui, ou seja, a tensão
a qual a coluna de água está submetida aumenta.
Em decorrência
da evaporação da água ao nível de
paredes celulares no mesófilo e da redução
localizada do potencial hídrico (Figura 14), há
o deslocamento de água a partir do interior das células.
Está redução do "status" energético
da água, que se transfere de célula para célula
favorece o deslocamento da água dos vasos do xilema,
seguindo um gradiente decrescente de potencial hídrico.
O "continuum" de moléculas de água,
formando uma corrente transpiratória somente é
possível em função da forte ligação
(coesão) entre as moléculas de água por
pontes de hidrogênio causando uma tensão.
A saída da água dos vasos do xilema em quantidades
maiores do que a reposta, induz no seu interior o surgimento
de uma tensão que se transmite através de toda
a extensão longitudinal do sistema lenhoso até
as raízes (Figura 15). Sempre que o potencial hídrico
do interior das raízes for mais baixo do que o do solo,
se verifica o processo de absorção de água,
considerando-se que a disponibilidade hídrica e a condutividade
hidráulica do solo permitam a sua ocorrência.
Figura 15.
Esquema mostrando a origem das tensões (ou seja, pressão
negativa) na água presente na parede celular de uma célula
vegetal. Com a evaporação da água que forme
uma película sobre a parede, meniscos são formados
nos interstícios da parede, os quais estão sob
tensão. Quando o raio de curvatura diminui, a pressão
diminui, ou seja, a tensão a qual a coluna de água
está submetida aumenta.
A ascenção
da água ocorre em virtude da grandeza da força
motriz (w)
normalmente desenvolvida pela transpiração, esta
se constitui na força mais importante de absorção
da água por parte das plantas, tanto em volume de água
como em termos de velocidade de translocação.